Litter fall inputs and litter decomposition represent important components in the nutrient cycling processes of forest ecosystems because the turnover of litter is a major pathway by which nutrients are transferred to forest soils (Bray and Gorham 1964, Ribeiro et al. 2002, Berg and Laskowski 2006). However, the quantity and quality of litter fall and release of nutrients from decomposing litter depend on several ecological factors and forest management activities, such as tree species, climate, site quality, stand age, stand density, fertilization, and thinning (Berg and Laskowski 2006, Inagaki et al. 2008, Kim et al. 2013).
An understanding of how litter fall quantity and litter decomposition changes with basal area in forest ecosystems is important to attain because the amount of basal area can alter litter fall flux, litter decomposition, and nutrient cycling processes in conjunction with changes in site environmental factors (Prescott et al. 2004, Berg and Laskowski 2006). In addition, litter fall fluxes tend to decrease roughly in proportion to the amount of basal area (Berg and Laskowski 2006, Inagaki et al. 2008, Kim et al. 2009), while litter decomposition rates were increased (Piene and Van Cleve 1978), not changed (Kim et al. 1996), or decreased (Blanco et al. 2011) with decreased basal area. The dynamics of litter fall and litter decomposition rates can also be influenced by the intensity of canopy removal at various levels of basal area because of the change in abiotic factors, such as temperature, moisture and soil properties, and by biotic factors such as nitrogen and lignin concentration and the abundance of microbial communities (Prescott et al. 2004, Berg and Laskowski 2006, Davidson and Janssens 2006, Blanco et al. 2011).
Although the dynamics of litter fall and litter decomposition induced by canopy removal have received considerable research attention (Kim et al. 1996, Prescott et al. 2004, Inagaki et al. 2008, Kim et al. 2012 a, b), there is a paucity of information about the qualitative and quantitative changes in litter fall and nutrient dynamics at various levels of basal area. The objectives of this study were 1) to measure litter fall and nutrient (C, N, P, K, Ca, Mg) fluxes, and 2) to determine patterns of nutrient release from decomposing needle litter at various levels of basal area in a red pine stand.
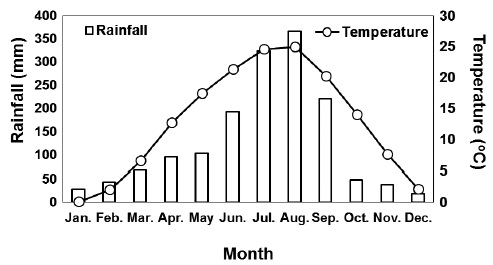
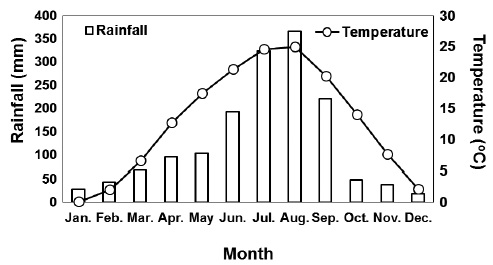
The study was conducted in the Hwangmaesan mountain forest, which is located in Sancheong-gun, Gyeongsangnam-do, Korea (Fig. 1). The mean monthly precipitation and temperature over the 30 years (1981-2010) from Sancheong meteorological observation network (35° 24′ N, 127 52′ E) near the study site were shown in Fig. 1 (KMA 2011). The soil in the study site is a well-drained, slightly wet, dark brown forest soil originating from granite, with a loamy texture and pH of 4.6-5.1. The study site consisted of an approximately 40-year-old natural red pine pure stand.
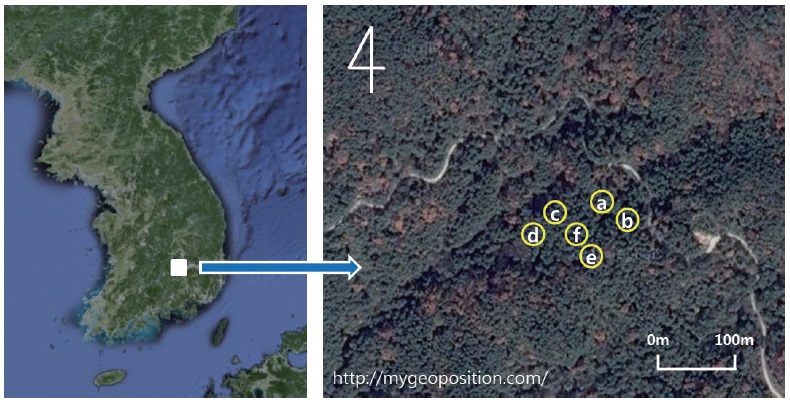
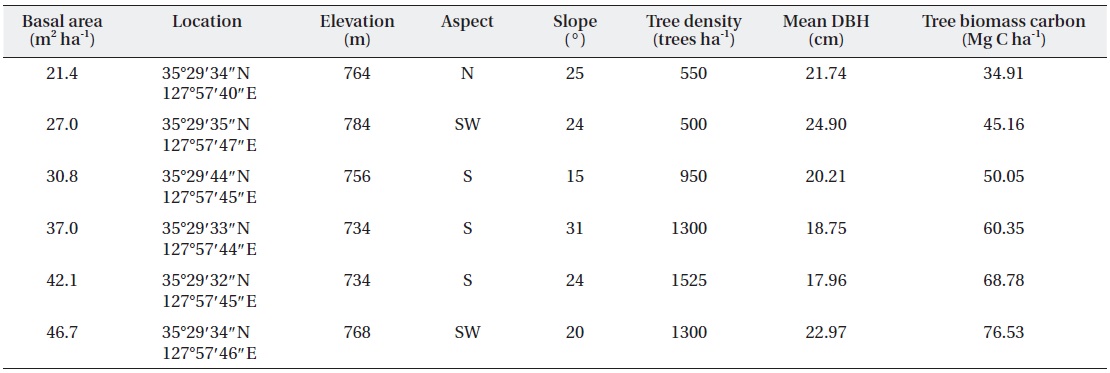
The experimental design consisted of six 20 m × 20 m plots at various levels of basal area in adjacent, natural red pine stands a small distance apart (< 20 m) under similar environmental conditions to minimize spatial variations in the soil properties (Fig. 2). The mean tree density ranged from 500 trees ha-1 to 1,525 trees ha-1, and the basal area ranged approximately from 21.4 m2 ha-1 to 46.5 m2 ha-1 (Table 1). The variation in tree density and basal area was due to forest-tending works conducted in 2005, which is the most common forest management practice in Korea (Kim et al. 2009).
[Table 1.] Stand characteristics at various levels of basal area in a red pine stands
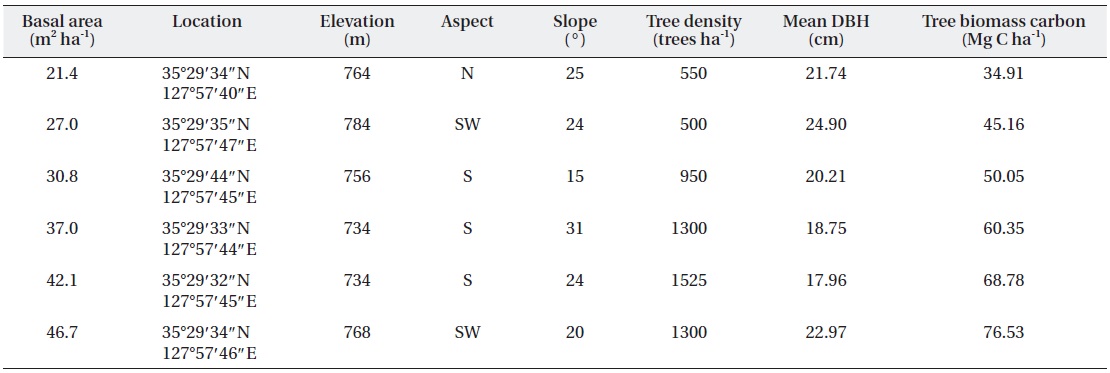
Stand characteristics at various levels of basal area in a red pine stands
To measure litter fall fluxes at various levels of basal area, five circular litter traps with a surface area of 0.25 m2 were installed 60 cm above the forest floor at each plot on 10 December 2007. Litter was collected at monthly intervals between January and December 2008. The needle litter from each trap was transported to a laboratory and then oven-dried at 65℃ for 48 h. All dried samples were separated into needle, bark, cones, branches, and miscellaneous components, and each portion was weighed. The needle litter samples were ground in a Wiley mill to pass through a 40-mesh (0.425 mm) stainless steel sieve. The carbon (C) and nitrogen (N) concentrations from the ground materials were determined with an elemental analyzer (CE Instruments EA1110, ThermoQuest Italia S.P.A. Italy). The other nutrient concentrations (P, K, Ca, Mg) were analyzed by ICP (Perkin Elmer Optima 5300, USA) with a standard method (National Institute of Agricultural Science and Technology 2000). Nutrient fluxes due to needle litter fall were calculated by multiplying needle litter fall mass by nutrient concentrations.
The needle litter decomposition rates at various levels of basal area were measured using the litterbag technique. Fresh needle litter from each treatment was collected from the forest floor during late November 2007. After collection, the litter was air-dried at room temperature for 14 days and a sample of approximately 10 g was weighed to the nearest 0.01 g and placed in a 30 cm × 30 cm nylon net bag with a mesh size of 0.1 mm. Subsamples from the litter were also taken to determine the oven dried mass after heating at 65℃ for 48 h. Total 90 litterbags (6 basal area treatments × 3 time collection × 5 replications) were randomly placed on the forest floor in 10 December 2007. The litterbags were held fast by 10-cm-long metal pins and collected three times (21st March, 23rd July, 11th December 2008) from each plot for one year. After collection, each litterbag sample was oven-dried at 65℃ for 48 h, weighed, and the mass loss rates determined. The needle litter samples collected for a litter bag were ground in a Wiley mill to pass through a 40 mesh (0.425 mm) stainless steel sieve. Nutrient concentrations from the ground materials were determined on an elemental analyzer and analyzed by ICP.
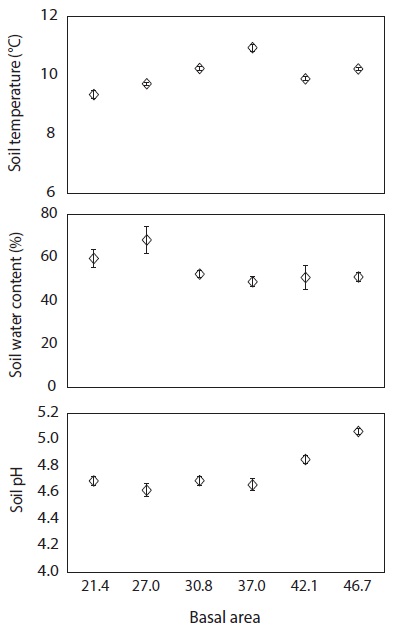
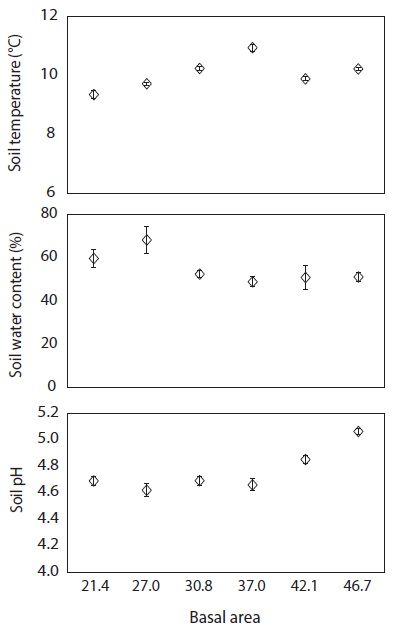
To determine soil environmental factors at various levels of basal area, soil temperature from five points within each plot adjacent to the litter decomposition bags was measured monthly at 20 cm depth using a digital soil temperature probe. In addition, soil core samples were collected at 20 cm depth using an Oakfield sampler. The soil samples were placed in plastic bags, transported to a laboratory and dried in an oven for 48 h at 105℃ to quantify the soil gravimetric water content and the soil pH (in 1:5 soil to water suspension) by glass electrode. All data were analyzed by using Pearson’s correlation at
>
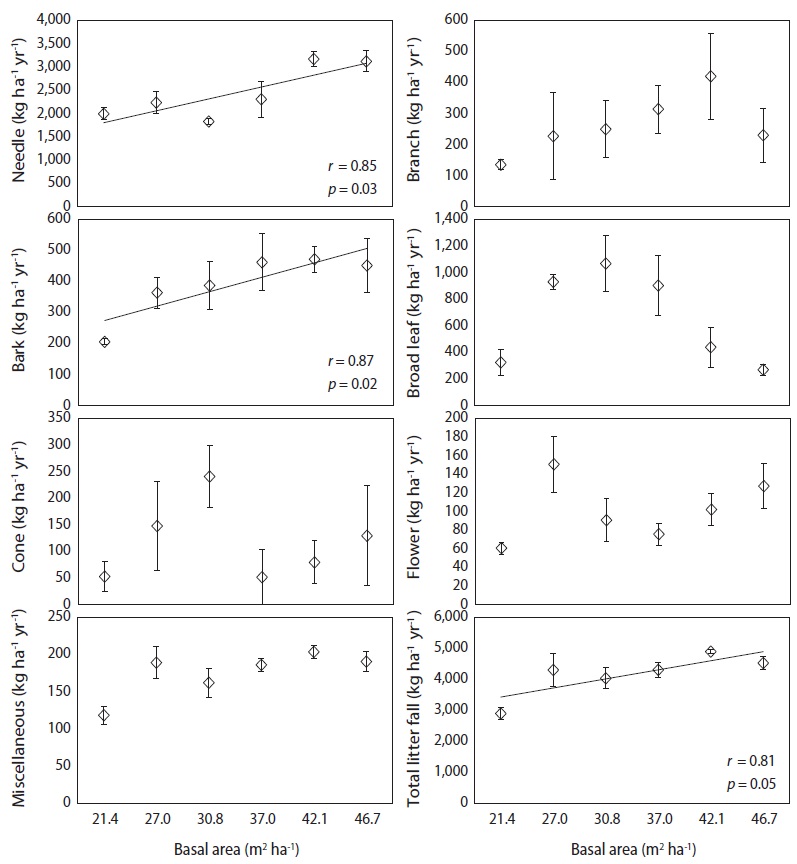
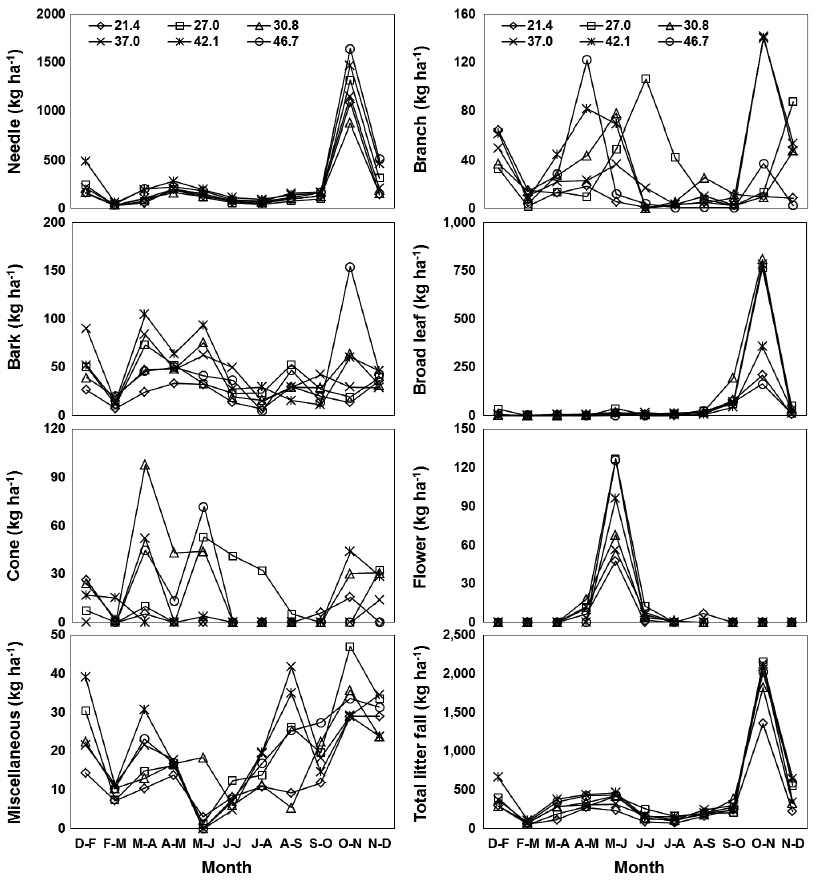
Litter fall and nutrient fluxes via needle litter at various levels of basal area
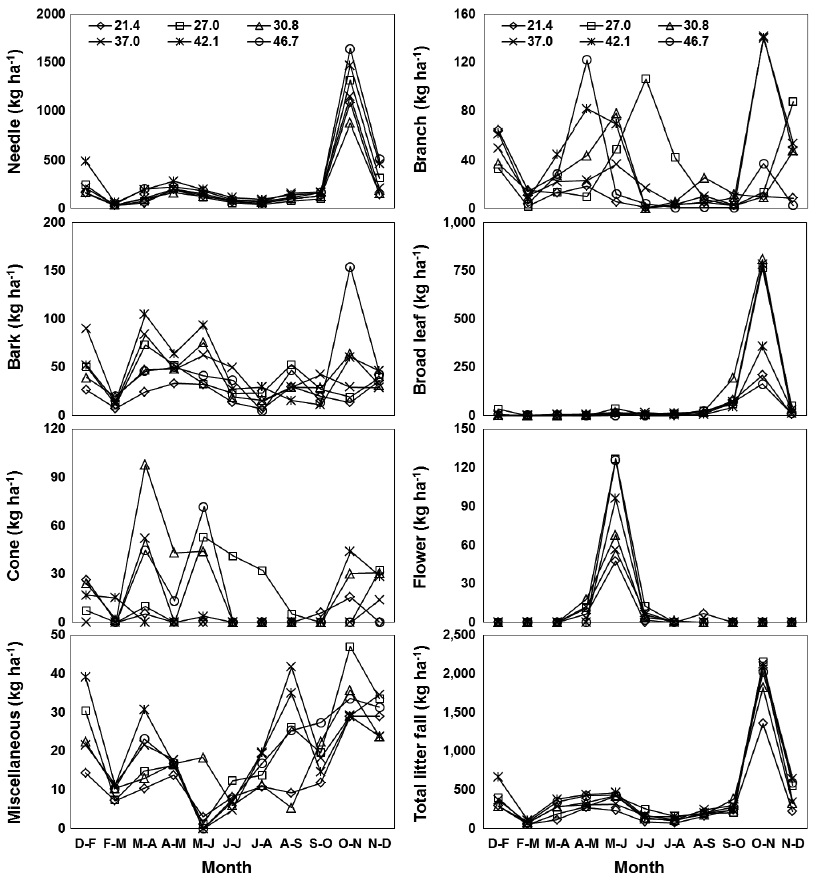
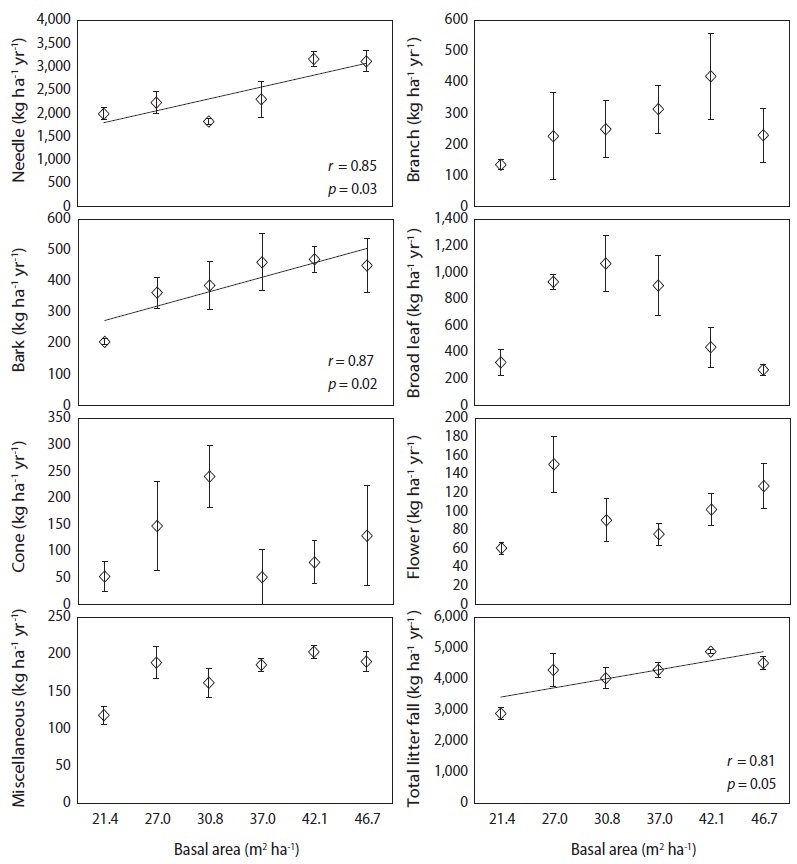
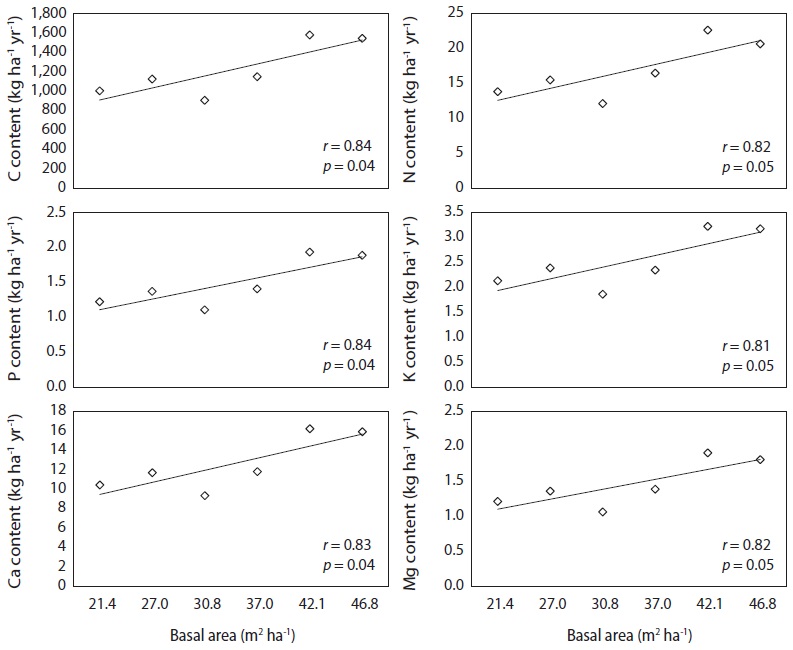
Monthly needle litter fluxes were higher in the 46.7 m3 ha-1 plot than in the plots with other basal areas during the months of heavy litter fall (Oct. to Nov., 2008), while needle litter fluxes in other months were not affected by the basal area treatments (Fig. 3). Monthly patterns of woody litter such as branch, bark, or cone litter were irregular in all basal treatments. A significant positive correlation was found between the increase in basal area and annual fluxes of the needle, bark, and total litter fall, but those of the broad leaf, cone, flower and miscellaneous litter were not significantly correlated (
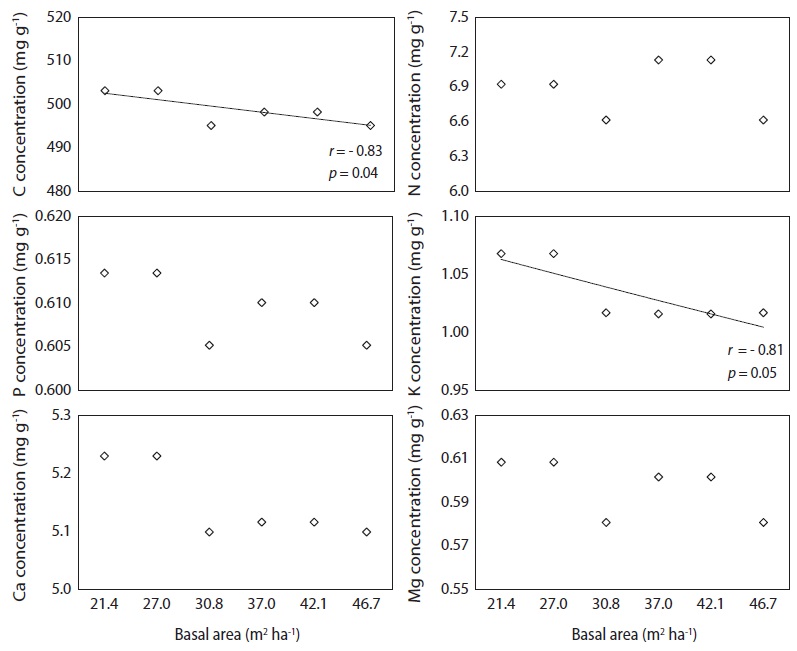
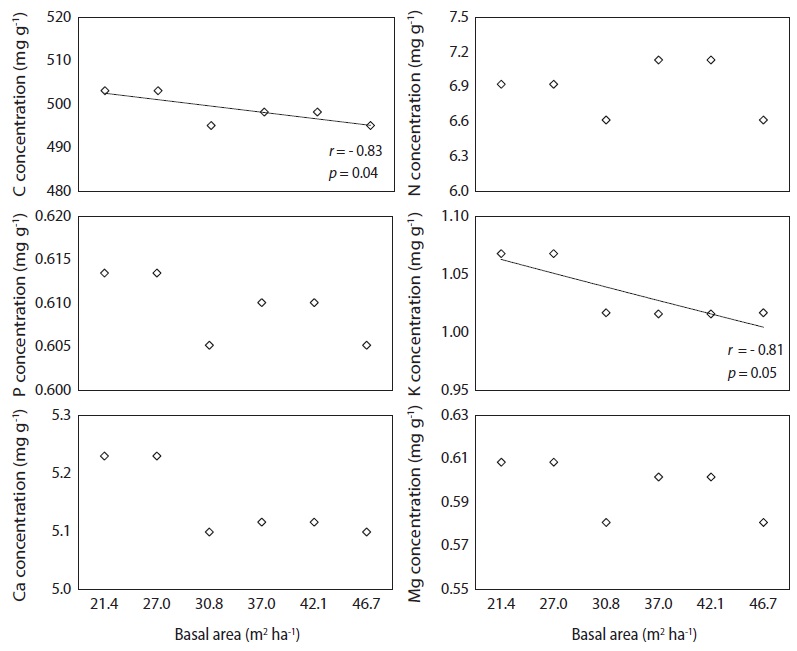
Carbon and K concentration in needle litter were negatively correlated with basal area (Fig. 5), while basal area was not correlatd with N, P, Ca, and Mg concentrations. There was a positive correlation (
>
Nutrient dynamics from decomposing needle litter
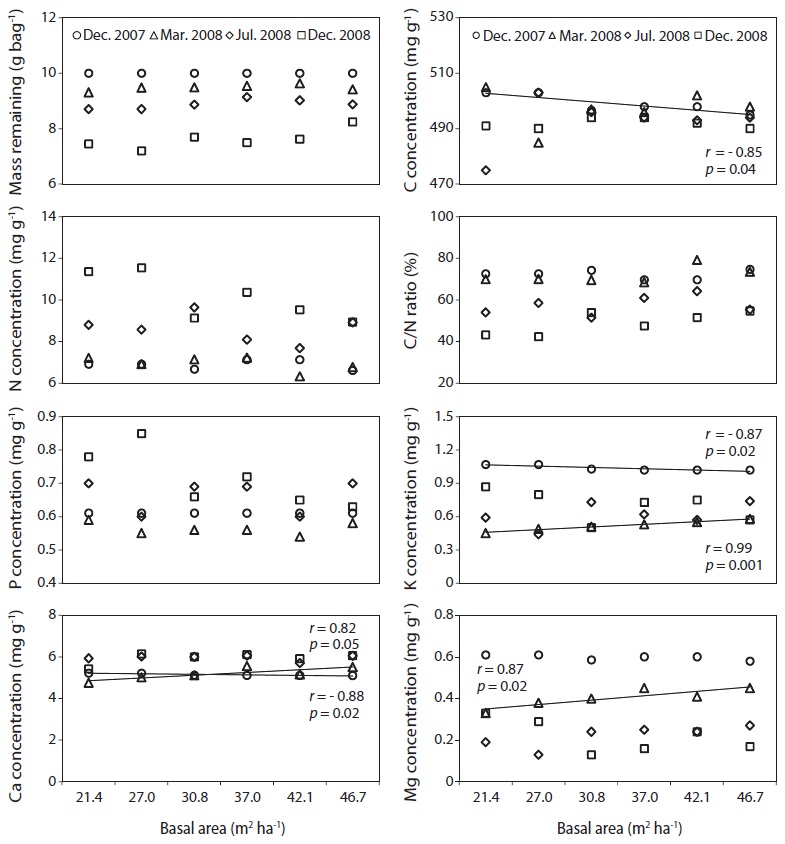
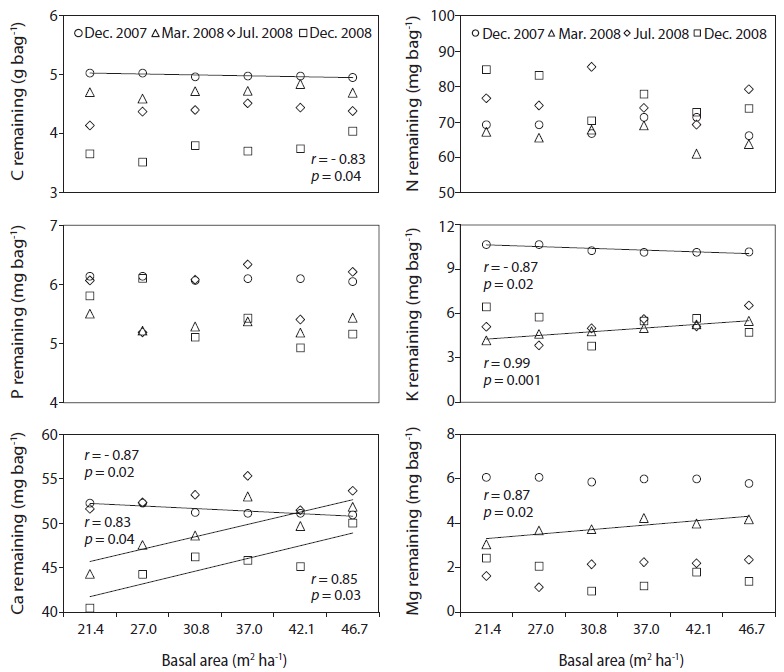
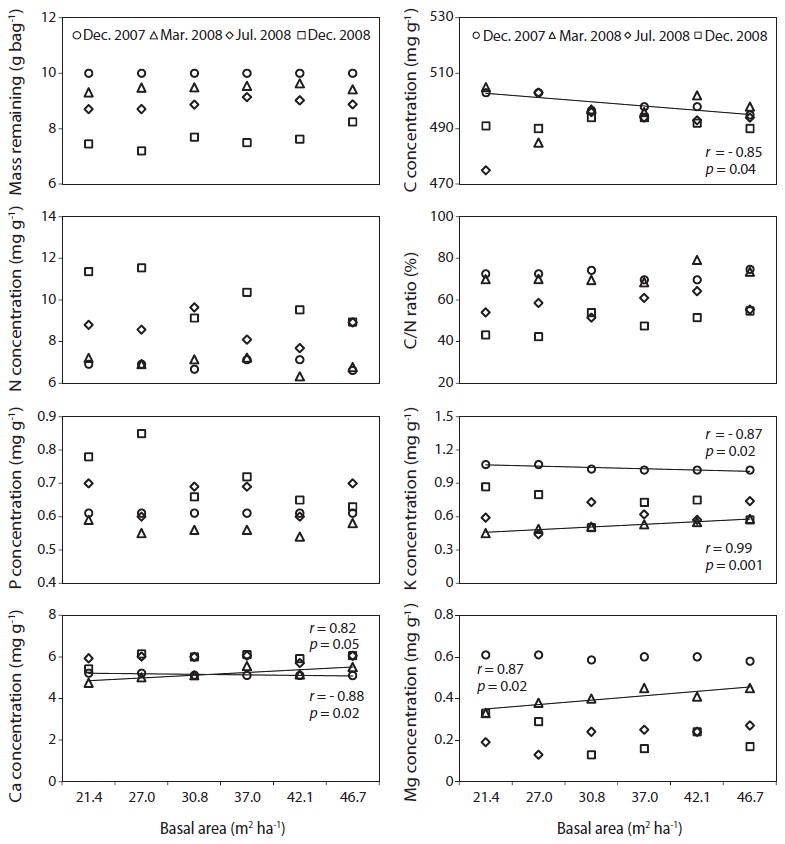
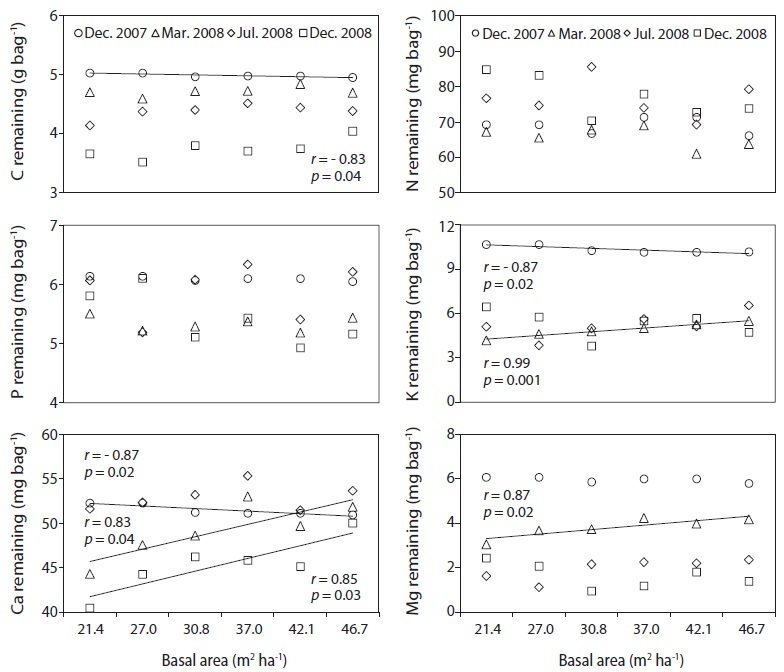
The C, N and P concentrations (mg g-1) and stocks remaining (g bag-1) in decomposing needle litter were not correlated with the basal area during the study period (Fig. 7, 8). Additionally, abiotic factors such as soil temperature, soil water and soil pH were not correlated with the various levels of basal area over the study period (Fig. 9). In all plots, C from decomposing needle litter was released over the decomposition process, while N was immobilized. In contrast to C, P from decomposing needle litter was released in the initial three months, but immobilized in the late stage of decomposition. The decomposing needle litter in this study showed no significant variation in nutrient (C, N, and P) stocks remaining among the treatment plot. However, the concentration and stocks remaining of K, Ca, and Mg were positively correlated with increased basal area treatments during the initial three months of incubation.
>
Litter fall and nutrient fluxes via needle litter at various levels of basal area
Litter fall fluxes in all of the basal area treatments followed similar monthly patterns because litter fall fluxes are affected by weather patterns (Jeong et al. 2009, Kim et al. 2012b). A strong monthly peak pattern of needle, broad leaf, flower and total litter fall occurred in all basal area treatments. The patterns for total litter fall closely reflected the monthly patterns of needle litter fall, which is a major component of litter fall at most levels of basal area in a red pine stand.
Annual fluxes of needle, bark and total litter fall linearly increased with increased basal area because tree removal was highest in plots with the lower basal area treatments. Similarly, basal area was generally the major factor limiting litter fall amount in many temperate forest ecosystems (Bray and Gorham 1964, Inagaki et al. 2008, Kim et al. 2012b). However, annual fluxes of branch, cone, and flower litter were not related to the levels of basal area, possibly because these components may have been affected by climatic factors such as storms or strong winds with high spatial variation (Christensen 1975). The fluxes of total litter fall ranged from 2910 kg ha-1 yr-1 in the plot with lowest basal area to 4528 kg ha-1 yr-1 in the plot with highest basal area. This value was lower than the 5475 kg ha-1 yr-1 observed in red pine forests in southern Korea (Jeong et al. 2009), and the global mean value of approximately 5500 kg ha-1yr-1 for warm temperate forests (Bray and Gorham 1964). The annual litter fall fluxes in this study were expected to decrease in response to canopy removal after forest-tending work, although this reduction effect on litter fall fluxes had disappeared several years after thinning in other pine stands (Roig et al. 2005, Blanco et al. 2006).
Nutrient concentrations in needle litter varied considerably and were influenced by basal area treatments. The low C and K concentrations in needle litter derived from the plots with higher basal area could be due to the nutrient dilution effect by high needle litter production. However, basal area was a poor indicator of N, P, Ca, and Mg concentrations in needle litter because the variation in the nutrient concentration of needle litter depends upon the combined effect of soil nutrient status, stand growth, leaching loss via throughfall, and resorption potential during needle senescence (Kim et al. 2013). In contrast, the N concentration of needle litter in
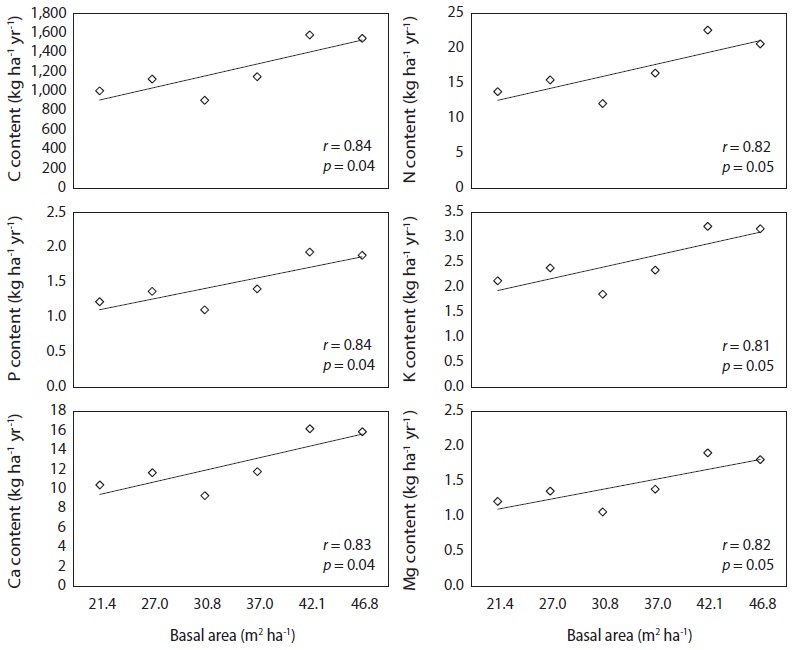
Variation in nutrient fluxes by needle litter fall with basal area was attributed to the differences in needle litter fluxes rather than in the nutrient concentration of the needle litter. The nutrient fluxes in this study matched the values observed in other red pine stands, which were 1.70 Mg C ha-1 yr-1, 17.4 kg N ha-1 yr-1, 1.5 kg P ha-1 yr-1, and 19.4 kg Ca ha-1 yr-1 (Kim et al. 2012b).
>
Nutrient dynamics from decomposing needle litter
The trends of mass remaining from decomposing needle litter at various levels of basal area could be attributed to variations in resource availability (e.g., light, temperature, water, and nutrients) because of the similar substrate quality, such as N concentration (Fig. 5) and C/N ratio (Fig. 7) of needle litter in the study sites. In addition, the lack of a significant relationship between mass remaining and basal area in this study could be due to the lack of any significant change in resource availability at the various levels of basal area (Fig. 9). For example, abiotic factors such as mean annual soil temperature, soil water content, and soil pH, which are regarded as the main factors influencing litter decomposition (Prescott et al. 2004), were not correlated with basal area treatments. Similarly, partial canopy removal in coniferous forests did not increase the rates of needle litter decomposition due to complex ecosystem-level responses following canopy removal (Kim et al. 1996). Also, reduction of litter decomposition rates in low basal area
Carbon, N, and P concentration (mg g-1) and stocks remaining (g bag-1) from decomposing needle litter showed no dependence on basal area, although K, Ca, and Mg concentration and stocks remaining were affected by increased basal area, with increased nutrient loss in the low basal area plots during the initial incubation period (Mar. 2008). The decreased K, Ca, and Mg concentration and stocks remaining in the low basal area plots could have been subjected to increased leaching loss by throughfall under increased canopy gaps compared with the high basal area plots (Tietema and Wessel 1994). However, the lack of any significant relationship in N concentration and stocks remaining from decomposing needle litter at various levels of basal area could be due to the combined effect of responses following canopy removal such as reduced solubility (Schreeg et al. 2013), immobilization of throughfall inorganic N (Van Vuuren and Van der Eerden 1992), and microbial N mineralization during the decomposition process (Tietema and Wessel 1994). The nutrient concentration and stocks remaining from decomposing needle litter are consistent with other pine studies in all basal area treatments (Kim et al. 2012a).
In summary, the quality and quantity of nutrient fluxes and concentrations associated with litter fall components in a red pine stand were greatly altered at various levels of basal area. The nutrient fluxes by needle litter fall were positively correlated with increased basal area, whereas the concentrations were not significantly affected by basal area except for C and K. Mass loss rates and C, N, and P stocks remaining from decomposing needle litter showed no relationship with basal area, whereas the K, Ca, and Mg concentration and stocks remaining were positively correlated with increased basal area during the initial stage of decomposition. The results indicate that nutrient dynamics through litter fall and litter decomposition could vary considerably with the different levels of stand basal areas in a red pine stand.